Gastrointestinal smooth muscles
乮徚壔娗暯妸嬝乯偲栻暔
丂
1. 徚壔娗暯妸嬝
嵶挿偄朼悗宍傪帵偡嵶朎偑屳偄偵暯峴偵暲傫偱扨埵偲側傞彫偝側懇傪嶌傞丅嵶朎撪偵偼傾僋僠儞偲儈僆僔儞偑
懚嵼偡傞偑丄惍慠偲暲傫偱偄傞偙偲偼側偄丅偟偽偟偽傾僋僠儞偲寢崌偟偰dense
body乮擹枾懱乯偑擣傔傜傟丄
偙傟偑崪奿嬝偱偺Z-band
偺婡擻傪壥偨偟偰偄傞偲峫偊傜傟偰偄傞丅傑偨丄彫朎懱偺敪払傕崪奿嬝偲斾妑偟偰
挊偟偔彮側偔丄嬝尨慄堐偵増偭偰暲傇偙偲偼側偄丅嵶朎枌昞柺偵偼暯妸嬝偵摿挜揑側caveole乮昞柺彫鈢乯偑
悢懡偔懚嵼偡傞丅
崪奿嬝偲偺斾妑
|
|
暯妸嬝
|
崪奿嬝
|
|
宍懺
|
朼悗宍偱懇傪側偡丅
|
墌摏宍
|
|
壜擻側廂弅乮尦偺挿偝偺乯
|
1/4
|
1/2
|
|
傾僋僠儞偲儈僔儞偺斾
|
10-20丗1
|
2丗1
|
|
嬝彫朎懱偺敪払
|
彮側偄丅
|
懡偔偺彫朎懱偑惍慠偲嬝尨慇堐偵
増偭偰暲傇丅
|
丂
|
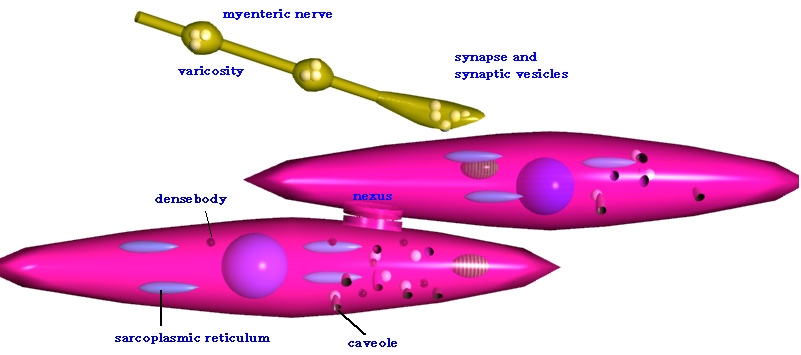
暯妸嬝偺摿挜傪昞偡奣棯恾
崪奿嬝偺傛偆偵嬝尨慄堐偑惍慠偲懡悢暲傫偱偄傞偙偲側偄丅暯妸嬝嵶朎偼帺棩恄宱偁傞偄偼枛徑恄宱偺巟攝傪丄昁偢偟傕捈愙庴偗偰偄側偄丅懇偵側偭偨暯妸嬝嵶朎偺娫傪憱峴偡傞恄宱偼丄懡偔偺varicosity乮恄宱朿棽晹乯偲廔枛偵傛傝丄暯妸嬝嵶朎偵壔妛揱払傪峴偆偑丄昁偢偟傕柧椖側恄宱嬝愙崌晹傪宍惉偟側偄丅捈愙丄恄宱偲愙崌晹傪宍惉偟側偄暯妸嬝嵶朎偼丄嵶朎偳偆偟偑gap
junction乮nexus乯偵傛傝揹婥揑偵宷偑傝丄扙暘嬌偑師偺嵶朎傊揱傢傞丅nexus偼杦偳偺暯妸嬝偱
尒傜傟傞丅
|
2. 恄宱巟攝偲恄宱丒暯妸嬝揱払
徚壔娗塣摦偼奜棃恄宱偲撪嵼恄宱偵傛傝惂屼偝傟偰偄傞丅奜棃恄宱偲偟偰丄岎姶恄宱偲暃岎姶恄宱偑巟攝偟偰偄傞偑丄
偙傟傜奜棃恄宱偺悢偼撪嵼恄宱偵斾傋偰嬌傔偰彮側偄丅挵恄宱憄偵偼傾儈儞丄傾儈僲巁丄儁僾僠僪宯側偳偺拞悤恄宱
偵懚嵼偡傞傎傏慡偰偺恄宱揱払暔幙傪娷傓撪嵼恄宱偑懚嵼偡傞丅廬偭偰徚壔娗塣摦偼撪嵼恄宱偺妶摦偵傛傝庡偵惂屼偝傟
偰偄傞偲尵偊傞丅
堦曽丄徚壔娗偱偼僇僴乕儖乮Cajal乯偺娫幙嵶朎傗堦晹偺暯妸嬝偑帺敪惈偵妶摦揹埵傪敪惗偟丄徚壔娗塣摦傪惗偠偰偄傞丅
揈弌慻怐偱傕帺敪塣摦傪婰榐偡傞偙偲偑偱偒傞偺偼偙偺偨傔偱偁傞丅
暯妸嬝偼崪奿嬝偵斾傋斾妑揑抶偄廂弅傪偡傞丅暯妸嬝廂弅傪惗偠傞 Ca2+
偵傛傞僔僌僫儖揱払偵偼丄嵶朎奜偲嵶朎撪
僗僩傾乕偺椉曽偺 Ca2+ 偑梡偄傜傟丄 Ca2+
棙梡偺僷僞乕儞偼崪奿嬝傗怱嬝偲堎側傞丅
倎丄奜棃恄宱偲撪嵼恄宱
暃岎姶恄宱宯偼柪憱恄宱偑巟攝偟偰偄傞偑丄壓晹寢挵傗捈挵偱偼愬晹暃岎姶恄宱傕巟攝偟偰偄傞丅偟偐偟丄椺偊偽僱僐
偱偺柪憱恄宱墦怱惈慄堐偺悢偼彮側偔丄徚壔娗恄宱憄嵶朎偺栺堦枩暘偺堦偲尵傢傟傞丅帠幚丄暯妸嬝嵶朎偵捈愙廔傢傞
恄宱慄堐偼彮側偔徚壔娗暯妸嬝慡懱傪巟攝偡傞偙偲偼側偄丅撪嵼僐儕儞恄宱傪巋寖偡傞偲暯妸嬝嵶朎偱嫽暠惈愙崌晹揹埵
(excitatory junction potentials,
e.j.ps)偑偲傟傞丅堦曽丄岎姶恄宱宯偲偟偰丄壓晹挵娫枌恄宱愡偐傜愡屻慄堐偑偺傃偰
偄傞偑丄杦偳偑撪嵼恄宱憄偵僔僫僾僗傪嶌偭偰廔傢傞丅廬偭偰丄岎姶恄宱傪巋寖偟偰傕暯妸嬝嵶朎偱梷惂惈愙崌晹揹埵
(inhibitory junction potentials, i.j.ps)
偑偲傟傞偙偲偼側偄丅恄宱憄撪偵偍偄偰 cholinergic neuron
偺恄宱廔枛
偵廔傢傝丄僔僫僾僗慜梷惂偵傛傝丄ACh
梀棧傪梷惂偟偰撪嵼恄宱偺揱払傪梷惂偟偰偄傞丅
暯妸嬝嵶朎偵梷惂惈偺揹埵傪敪惗偝偣傞揱払偼丄旕傾僪儗僫儕儞旕僐儕儞(Non-adrenergic
non-cholinergic, NANC) 惈
揱払偲屇偽傟偰偄傞丅崪奿嬝偱偼懡偔偺庬傪偙偊偰塣摦恄宱偺恄宱揱払暔幙偼
acetylcholine 偱偁傝丄偙偺嫽暠惈巟攝偵
傛偭偰偺傒崪奿嬝偺嬞挘偑惂屼偝傟偰偄傞丅偙傟偵妑傋丄暯妸嬝偵偼梷惂惈恄宱巟攝偑懚嵼偟丄偦傟偑摦暔庬偵傛傝丄
挵娗晹埵偵傛傝挊偟偔堎側傞偙偲偑抦傜傟偰偄傞丅NANC 惈揱払暔幙偲偟偰
堦巁壔拏慺丄vasoactive intestinal peptide
(VIP), neurotensin, ATP
側偳偑堦晹偺徚壔娗晹埵偱摥偄偰偄傞偙偲偑曬崘偝傟偰偄傞偑丄埶慠偲偟偰梷惂惈恄宱偑枹抦偱
偁傞徚壔娗晹埵偑懡偔丄抩娚婡彉傪娷傔偰徻嵶偼抦傜傟偰偄側偄丅
倐丄帺敪塣摦
徚壔娗偺暯妸嬝嵶朎偵偼丄帺敪惈偵妶摦揹埵傪敪惗偡傞嵶朎丄巋寖偵傛偭偰妶摦揹埵傪敪惗偡傞嵶朎丄巋寖傪壛偊偰傕敪惗
偟側偄嵶朎偲偑偁傞丅堦晹偺暯妸嬝嵶朎偱敪惗偡傞帺敪惈偺揹埵偼 slow
wave 偲屇偽傟丄宍傗昿搙偼暯妸嬝偵傛偭偰
堎側傞偑丄悢 mV 偐傜悢廫 mV 偺怳暆偱 1-10 悢昩帩懕偡傞娚傗偐側揹埵偱
1 暘娫偵 1-20 屄敪惗偡傞丅扙暘嬌偑鑷抣偵
払偡傞偲妶摦揹埵傪敪惗偡傞丅堦晹丄儌儖儌僢僩寢挵昍傗惛娗側偳偱偼怱嬝偺傛偆側
spike 宆偺妶摦揹埵傪敪惗偡傞慻怐
傕偁傞丅tetrodotoxin 傗 atropine 偑柍岠偱偁傝slow wave
偼嬝尨惈偱偁傞丅擲枌嬝斅偲椫憱嬝憌偺娫偵懚嵼偡傞
Cajal cell 偐傜敪惗偟丄nexus
傪夘偟偰暯妸嬝慻怐慡懱偵揱傢傝挵塣摦傪惗偠傞丅妶摦揹埵偼儊僨傿僂儉偺
Na+ 傪彍嫀偟
偨傝丄tetrodotoxin偱 Na+
棳擖傪慾奞偟偰傕塭嬁傪庴偗側偄丅偟偐偟丄嵶朎奜偺 Ca2+
偵埶懚惈偑崅偔丄 Ca2+ 漢峈栻傗
Mn2+ 側偳偱 Ca2+ 棳擖傪梷偊傞偲妶摦揹埵偼弌側偔側傝丄暯妸嬝偺spike 偼
Ca2+ spike 偱偁傞偙偲偑暘偐傞丅
們丄暯妸嬝偺廂弅
乮1乯Ca2+ 偵傛傞僔僌僫儖揱払
抩娚帪偺暯妸嬝嵶朎撪偺 Ca2+ 擹搙偼60-200 nM 掱搙偱丄廂弅帪偼1-10 mM
偲側傞丅嵶朎奜塼偺 Ca2+ 擹搙偼 3mM 掱搙
側偺偱挊偟偄擹搙岡攝傪惗偠偰偄傞偑丄惷巭帪偺暯妸嬝嵶朎枌偼 Ca2+
偵懳偟杦偳摟夁惈偑側偄丅恄宱揱払暔幙偺摥偒偵傛傝丄
嵶朎撪 Ca2+
偑崅傑傞偲廂弅婡峔偑妶惈壔偝傟丄傾僋僠儞偲儈僆僔儞偺憡屳嶌梡偵傛傝廂弅傪惗偠傞丅
乮2乯嬝廂弅偵娭傢傞 Ca2+ 偺嫙媼尮
i) Ca2+ channel 傪捠偭偰偺奜偐傜偺 Ca2+ 棳擖
ACh 偑 muscarinic
庴梕懱偵寢崌偟偨帪側偳偱偼丄庴梕懱嶌摦惈偺旕慖戰惈梲僀僆儞 channel
偑奐偄偰 Ca2+ 偑奜偐傜
棳擖偡傞丅傑偨丄暯妸嬝嵶朎枌偑扙丂暘嬌偡傞偙偲偵傛傝丄揹埵埶懚惈偺
Ca2+ channel 偑奐偄偰 Ca2+ 偑棳擖偡傞丅
ii) 嵶朎撪 Ca2+ 僗僩傾乕偱偁傞 sarcoplasmic reticulum (SR) 偐傜偺
Ca2+ 曻弌
ACh 偑 muscarinic 偺 m1, m3, m5
庴梕懱偵寢崌偟偨帪側偳偱偼丄phospholipase C (PLC)
兝偺妶惈壔傪夘偟偰丄
inositoltrisphosphate (IP3) 偑嶻惗偝傟傞丅傑偨丄tyrosine kinase
偺妶惈壔偵傛傝 PLCg 偑 IP3 傪嶻惗偡傞丅
IP3 偑 SR 偵懚嵼偡傞 IP3 庴梕懱傪妶惈壔偟偰 Ca2+
傪曻弌偡傞丅IP3庴梕懱偼 Ca2+ 偺懚嵼壓偵偝傜偵妶惈壔偝傟傞丅
傑偨丄棳擖偟偨 Ca2+ 偑 SR 偵懚嵼偡傞 ryanodine 庴梕懱傪妶惈壔偟丄
Ca2+ 傪曻弌偡傞丅偙偺婡峔偼怱嬝偱偲偔偵尠挊偱丄
棳擖偟偨 Ca2+ 傛傝傕偼傞偐偵懡検偺 Ca2+ 傪 SR 偐傜曻弌偡傞丅
iii) 嵶朎幙偐傜偺 Ca2+ 偺攔弌梷惂
暯妸嬝嵶朎枌偺 plasma membrane Ca2+-ATPase 偺梷惂偵傛傞嵶朎奜傊偺
Ca2+ 攔弌梷惂丄偁傞偄偼 SR 偵懚嵼偡傞
SR Ca2+-ATPase (SERCA 2) 偺梷惂偵傛傞 SR 傊偺 Ca2+
庢傝崬傒梷惂偵傛偭偰傕嵶朎撪 Ca2+ 偼憹壛偡傞丅
乮3乯暯妸嬝偱偺嵶朎撪 Ca2+ 偺憹壛
徚壔娗暯妸嬝偱偼丄庴梕懱嶌摦惈 Ca2+ channel 偍傛傃揹埵埶懚惈 Ca2+
channel 偺椉曽偑摥偄偰偄傞丅傑偨丄IP3 庴梕懱
偍傛傃ryanodine 庴梕懱傪夘偟偰 Ca2+ 傪 SR
偐傜曻弌偡傞椉曽偺婡彉偑摥偄偰偄傞丅偦傟傜偺摥偒偺廳梫惈偼慻怐偲
傾僑僯僗僩偺庬椶偵傛傝堎側傞丅堦斒揑偵偼丄暯妸嬝廂弅偺憗偄戞堦憡偵偼僗僩傾乕偐傜偺
Ca2+ 偑丂棙梡偝傟丄抶偄戞擇憡
偵偼嵶朎奜偐傜偺Ca2+ 棳擖偑峷專偟偰偄傞丅
|
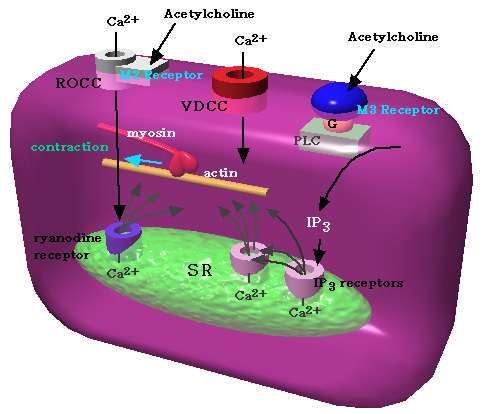
暯妸嬝嵶朎偱偺Ca2+摦堳婡峔
庴梕懱嶌摦惈乮ROCC乯偲揹埵埶懚惈Ca2+
channel傪捠偭偰奜偐傜偺棳擖偲IP3庴梕懱偲
ryanodine庴梕懱妶惈壔偵傛傞SR偐傜偺曻弌偑偁傞丅
|
丂
乮4乯儈僆僔儞偺儕儞巁壔偲廂弅
i) Ca2+ 僀僆儞偵傛傞儈僆僔儞寉嵔僉僫乕僛偺妶惈壔
憹壛偟偨 Ca2+
僀僆儞偼嵶朎撪僇儖儌僕儏儕儞偲寢崌偟丄儈僆僔儞寉嵔僉僫乕僛傪妶惈壔偡傞丅偦偺寢壥丄儈僆僔儞寉嵔偺
儕儞巁壔偑婲偙傝丄傾僋僠儞偲儈僆僔儞摢晹偱宍惉偝傟傞 cross bridge
偑夞揮偟偰廂弅偑惗偠傞丅廬偭偰丄廂弅偺嫮偝傪
寛傔傞偺偼堦媊揑偵偼丄儈僆僔儞寉嵔傪儕儞巁壔偡傞嵶朎撪 Ca2+
僀僆儞擹搙偲尵偊傞丅
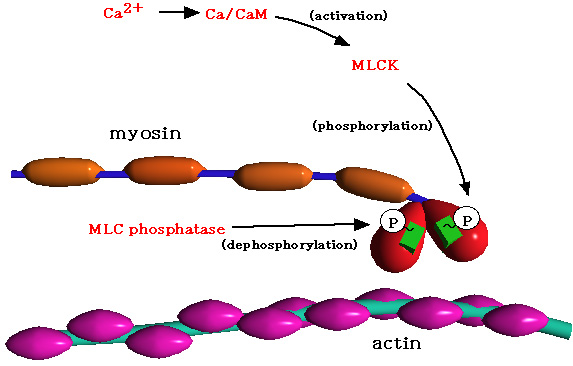
|
儈僔儞寉嵔偺儕儞巁壔偵傛傞cross
bridge偺夞揮
嵶朎撪偱崅傑偭偨Ca2+偼丄僇儖儌僕儏儕儞乮CaM乯偲寢崌偟丄MLCK傪妶惈壔偟丄儈僆僔儞寉嵔乮MLC乯偺摢晹傪
儕儞巁壔偡傞丅傑偨丄摢晹偵偁傞MLC-ATPase偑ATP傪暘夝偟丄cross
bridge偺夞揮偑惗偠傞丅
堦曽丄MLC
phosphatase偼丄扙儕儞巁壔斀墳傪扴偭偰偍傝丄kinase偲phosphatase偺僶儔儞僗偵傛傝丄
MLC偺儕儞巁壔検偑曐偨傟偰偄傞丅
|
ii) 儈僆僔儞寉嵔儕儞巁壔検偺挷愡
堦掕偺 Ca2+
僀僆儞擹搙壓偱丄儈僆僔儞寉嵔偺儕儞巁壔傪挷愡偡傞婡彉偑懚嵼偡傞丅暯妸嬝嵶朎枌偵懚嵼偡傞
m3 傗
兛1 庴梕懱偑妶惈壔偝傟傞偲丄掅暘巕検 GTP 寢崌抈敀幙偺傂偲偮偱偁傞
RhoA 偑妶惈壔偝傟丄RhoA kinase 傪夘偟偰
儈僆僔儞寉嵔僼僅僗僼傽僞乕僛偑儕儞巁壔偝傟丄妶惈偑梷惂偝傟傞丅偦偺寢壥儈僆僔儞寉嵔偺儕儞巁壔偑憹壛偟廂弅斀墳
偑弌傞丅偙傟偲偼暿偵丄PKC 偑 17kDa
抈敀傪夘偟偰丄傗偼傝儈僆僔儞寉嵔僼僅僗僼傽僞乕僛傪梷惂偡傞宯傕抦傜傟偰偄傞丅
偙傟傜偼丄Ca2+ 僀僆儞擹搙偑堦掕偱偁傞偺偵廂弅傪惗偠傞偺偱丄廂弅婡峔偺
乪Ca2+ 姶庴惈憹壛乫偲傕昞尰偝傟偰偄傞丅
偙偺傎偐丄嵶朎撪 cyclic GMP 偺憹壛偑 PKG
傪夘偟偰丄儈僆僔儞寉嵔僼僅僗僼傽僞乕僛傪妶惈壔偡傞丅
傑偨丄僇儖僔儏僂儉丒僇儖儌僡儏儕儞埶懚惈僾儘僥僀儞僉僫乕僛II偑儈僆僔儞寉嵔僉僫乕僛傪儕儞巁壔偟偰丄儈僆僔儞
寉嵔僉僫乕僛傪梷惂偡傞宯傕抦傜傟偰偄傞丅偙傟傜偺応崌偵偼丄Ca2+
僀僆儞擹搙偑堦掕偱偁傞偺偵抩娚傪惗偠傞偺偱丄
廂弅婡峔偺 乪Ca2+ 姶庴惈掅壓乫偲傕昞尰偝傟偰偄傞丅
倓丄暯妸嬝偺廂弅偲抩娚偺傑偲傔
暯妸嬝偼懱偺庡側拞嬻憻婍偵懚嵼偟偰偍傝丄徚壔婍丄寣娗丄銷泖丄巕媨側偳偵暘晍偟偰偄傞丅
偦偺惈幙偼婍姱偛偲偵堎側偭偰偍傝丄廂弅偲抩娚傪堷偒婲偙偡傾僑僯僗僩傕堎側偭偰偄傞丅
戝摦柆傗婥娗巟偺暯妸嬝偼妶摦揹埵傪敪惗偟側偄丅
廂弅宯
妶摦揹埵偼丄庡偲偟偰捈愙偺婡夿乮怢揥乯巋寖偲L宆揹埵埶懚惈Ca僠儍僱儖偵傛傝惗偠傞丅偙傟偑Ca僀僆儞偺
棳擖偺庡側宱楬偱偁傞丅傑偨丄懡偔偺暯妸嬝偵偼丄儕僈儞僪埶懚惈僀僆儞僠儍儞僱儖乮P俀X側偳乯偑偁傝丄帺棩恄宱
偐傜梀棧偝傟偨ATP側偳偵斀墳偟丄Ca僀僆儞偺棳擖傪堷偒婲偙偡丅偝傜偵丄偝傑偞傑側傾僑僯僗僩偵斀墳偡傞GPCR偑
懚嵼偟丄IP3嶻惗偲IP3庴梕懱巋寖傪夘偟偰丄SR偐傜Ca僀僆儞傪梀棧偡傞丅丂
丂丂丂
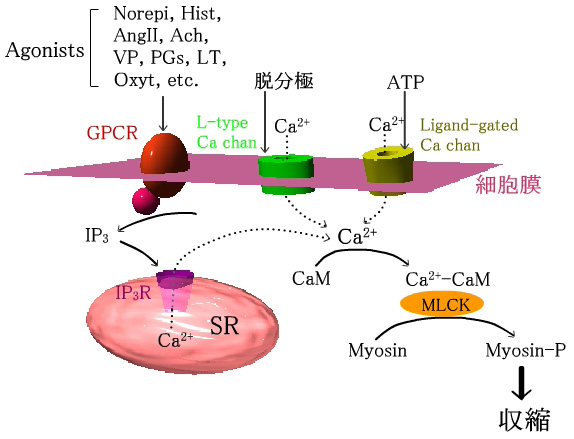 |
暯妸嬝偺廂弅宯
GPCR丗G抈敀嫟栶宆庴梕懱丄MLCK丗myosin light-chain kinase丄CaM丗
calmodulin丄SR丗嬝彫朎懱 |
抩娚宯
僇儕僂儉僠儍僱儖偲梷惂惈傾僑僯僗僩偵懳偡傞GPCR偼丄K僀僆儞棳擖丄們AMP嶻惗丄Ca僠儍儞僱儖偺
挷愡傪偟偰偄傞丅ANP庴梕懱偼枌寢崌宆GC傪丄婥懱偺NO偼壜梟惈GC傪巋寖偟丄們GMP偺嶻惗傪堷偒婲偙偡丅
PDE慾奞栻偼們AMP偲們GMP偺暘夝傪梷惂偡傞丅們AMP傗們GMP偼丄奺protein kinase傪夘偟丄MLCK偺儕儞巁壔傪
堷偒婲偙偡丅儕儞巁壔偝傟偨MLCK偼Ca2+-CaM偲偺恊榓惈偑掅壓偟抩娚傪堷偒婲偙偡丅
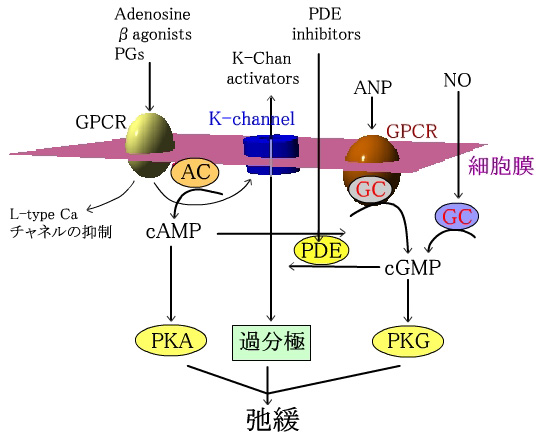 |
暯妸嬝偺抩娚宯
NO丗nitric oxide丄PKA丗protein kinase A丄PKG丗protein kinase G丄
PDE丗phosphodiesterase丄ANP丗atrial natriuretic peptide丄
AC丗adenylate cyclase丄GC丗guanylate cyclase
嶲徠丗儔儞僌丒僨乕儖乽栻棟妛乿乮2012乯 |
丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂丂敤丄嶰栘
 Download for Acrobat Reader
Download for Acrobat Reader
(2013/2/10)
丂
丂